

Анатомия и физиология сердечно-сосудистой и дыхательной систем у рептилий. Часть 2

Еще фото
Автор (ы): Л.А. Стоянов, врач ветеринарной медицины, начальник ветеринарно-медицинского отдела экзотических животных Международной ассоциации океанариумов и дельфинариев
Организация(и): Сеть океанариумов «Немо», г. Одесса
Журнал: №2 - 2013
Окончание. Начало в номере №1 за 2013 год
Благодарим редакцию журнала «Мир ветеринарии», Украина, за любезно предоставленную статью Л.А. Стоянова
Анатомия и физиология дыхательной системы
Знание нормальной анатомии и физиологии дыхательных систем рептилий крайне важно для врачебной практики. Хотя общее строение и функции легких у разных отрядов рептилий в целом одинаковы, у отдельных групп были найдены некоторые клинически значимые отличия. Недостаточное понимание анатомии и физиологии дыхательной системы рептилий не позволит эффективно лечить такие заболевания, как пневмония, и отрицательно скажется на умении обращаться с рептилией под действием анестезии.
Анатомия дыхательных путей
Дыхательные системы рептилий и млекопитающих сильно отличаются анатомически и физиологически. Существенные отличия наблюдаются также у разных отрядов рептилий и между видами внутри одного отряда, особенно при сравнении водных и наземных видов. Ниже приведены основные клинически значимые особенности анатомии дыхательных систем черепах, ящериц, крокодилов и змей.
Черепахи
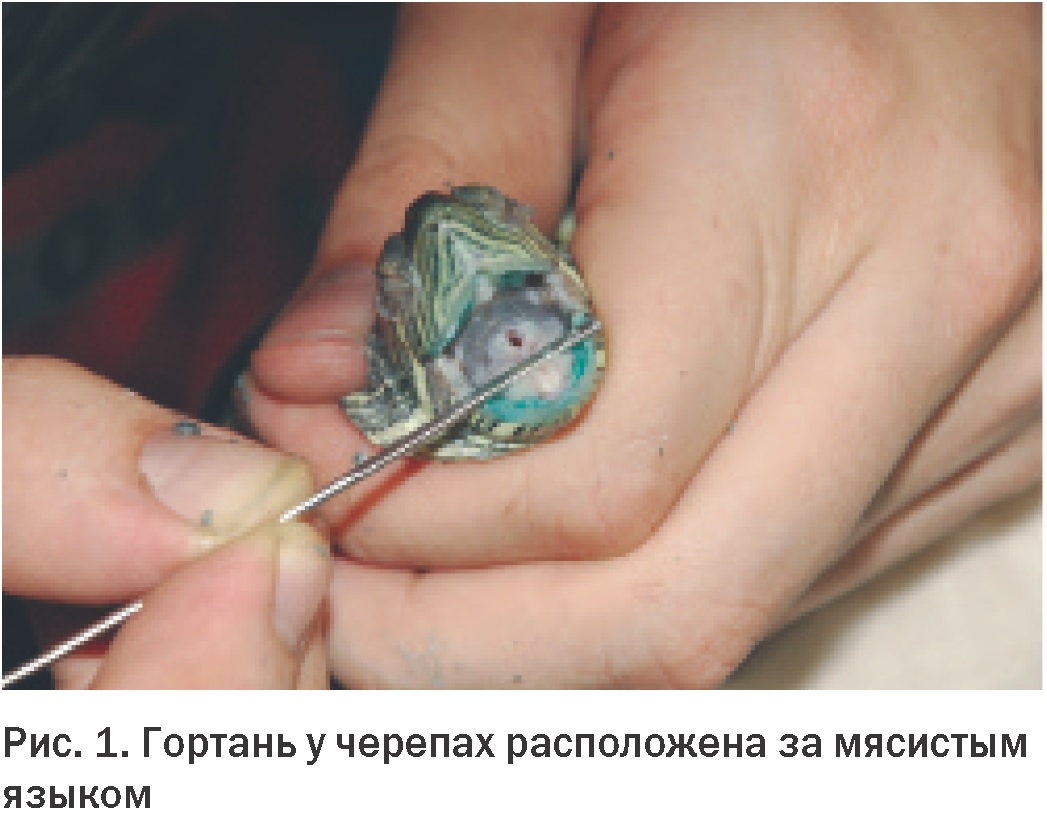
Гортань у черепах открывается у основания толстого мышечного языка в относительно каудальной части ротоглотки (Рис. 1). Такое расположение затрудняет доступ к нижним дыхательным путям через ротовую полость без применения наркоза. Трахея с полными хрящевыми кольцами раздваивается чуть ниже шеи. Далее парные бронхи в жесткой оболочке входят в легкое с дорсальной стороны.
Легкие черепах парные и мешковидные. Структуры, в которых происходит газообмен, названные фальвеолами, в противоположность альвеолам млекопитающих, открываются на окончаниях бронхов в воздушную полость легкого. При ближайшем рассмотрении структуры легкого обнаруживаются внутренние выступы и перегородки, напоминающие губку в поперечном разрезе. Общая анатомия нижних дыхательных путей черепах имеет большое значение. Воспалительный экссудат, в частности, образующийся при инфекционных заболеваниях, имеет свойство скапливаться в нижней части легкого, что препятствует его высвобождению через бронхи и трахею, как это происходит у млекопитающих.
У черепах прохождение газов через дыхательную поверхность для газообмена достигается различными путями. Эти различия зависят от вида, а также от того, водным или наземным он является. Как правило, и вдох, и выдох являются активными процессами. Следует, однако, отметить, что у водных видов выдыхание является более пассивным, тогда как у некоторых наземных видов более пассивным является вдох. Несмотря на то, что у черепах нет настоящей диафрагмы, многие из них обладают подобной ей мембранной структурой, которая частично разделяет грудную и брюшную полости и называется septum horizontale. У морских черепах это разделение отсутствует.
Как правило, у водных черепах в респираторном цикле работают четыре группы мышц. Сокращение диафрагмальных и поперечных мышц живота сжимает целомическую полость, что приводит к выдоху. Коракоидные и косые мышцы живота расширяют полость и обеспечивают вдох.
Интересно, что повторяющиеся горловые движения, которые владельцы животных часто принимают за дыхание, в действительности с ним не связаны и имеют обонятельную функцию. У сухопутных средиземноморских черепах (Testudo graeca) были найдены существенные отличия. У этого вида дыхательные движения обеспечивают мышцы плечевого пояса. Вдох происходит с сокращением зубчатых и косых мышц живота, а выдох – с сокращением поперечных мышц живота и грудных мышц.
Ящерицы
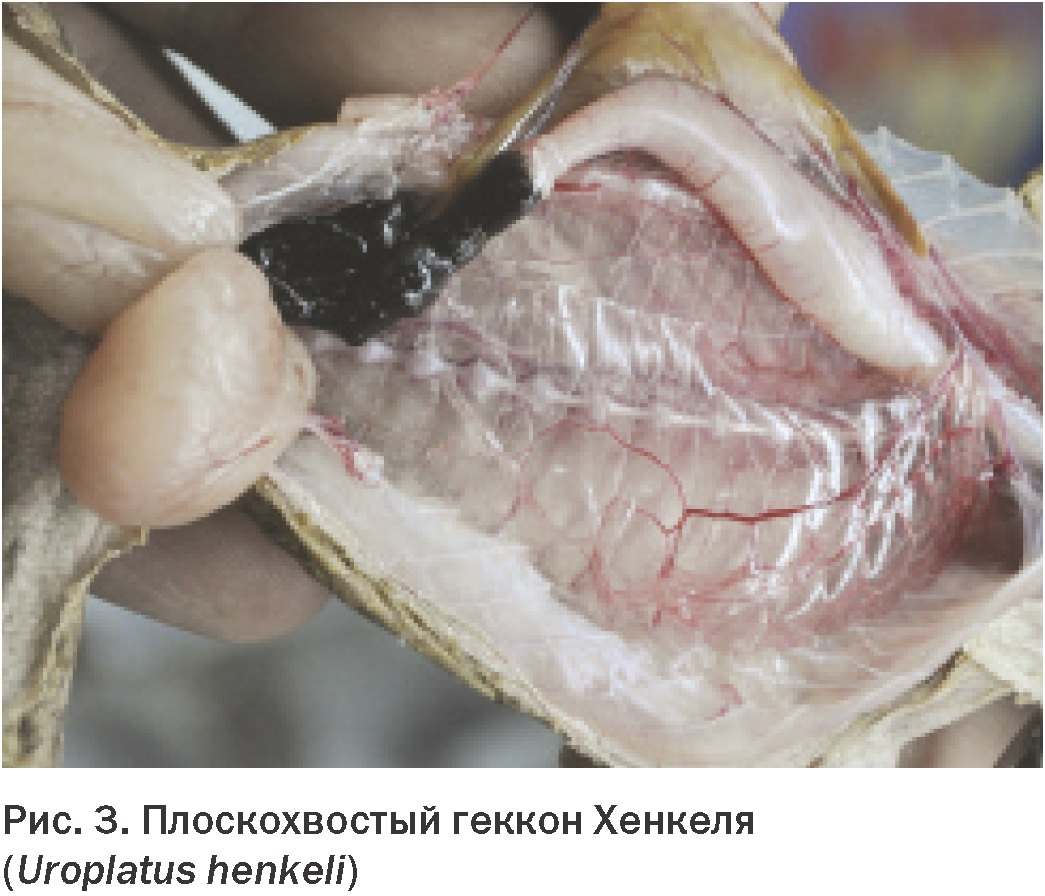
У ящериц расположение отверстия гортани в ротовой полости неодинаково (Рис. 2). У многих видов, в частности, у хищных, оно находится в ростральной части рта. У других – оно расположено вентральнее, у основания мясистого языка. У ящериц, как и у змей, кольца трахеи неполные. В отличие от черепах, их трахея не раздваивается в шейном отделе, а остается цельной до входа в грудную полость около сердца. Ниже трахея делится на левый и правый бронхи. Легкие ящериц примерно одинаковы по размеру и объему и их строение такое же, как у других рептилий (Рис. 3). У многих ящериц газообмен происходит в краниальной части легкого. Каудальное расширение легкого аналогично воздушному мешку у птиц. Эта особенность важна, так как продукты воспалительных процессов могут накапливаться в этой нереспираторной части легкого. В связи со слабой васкуляризацией этой области, инфекции могут удерживаться здесь и не поддаваться действию парентеральных медикаментов.
Как и у черепах, вдохи и выдохи у ящериц являются активными процессами. У большинства видов они совершаются при помощи межреберных мышц. Некоторые виды используют также гладкую мускулатуру внутри легкого для вдоха. У большинства видов сразу за вдохом и выдохом следует период задержки дыхания, различный по продолжительности. У некоторых ящериц, например, у варанов есть мембрана, отделяющая сердце и легкие от остальных внутренних органов, аналогично диафрагме между грудной и брюшной полостями у млекопитающих. Эта мембрана не имеет дыхательной функции.
Змеи
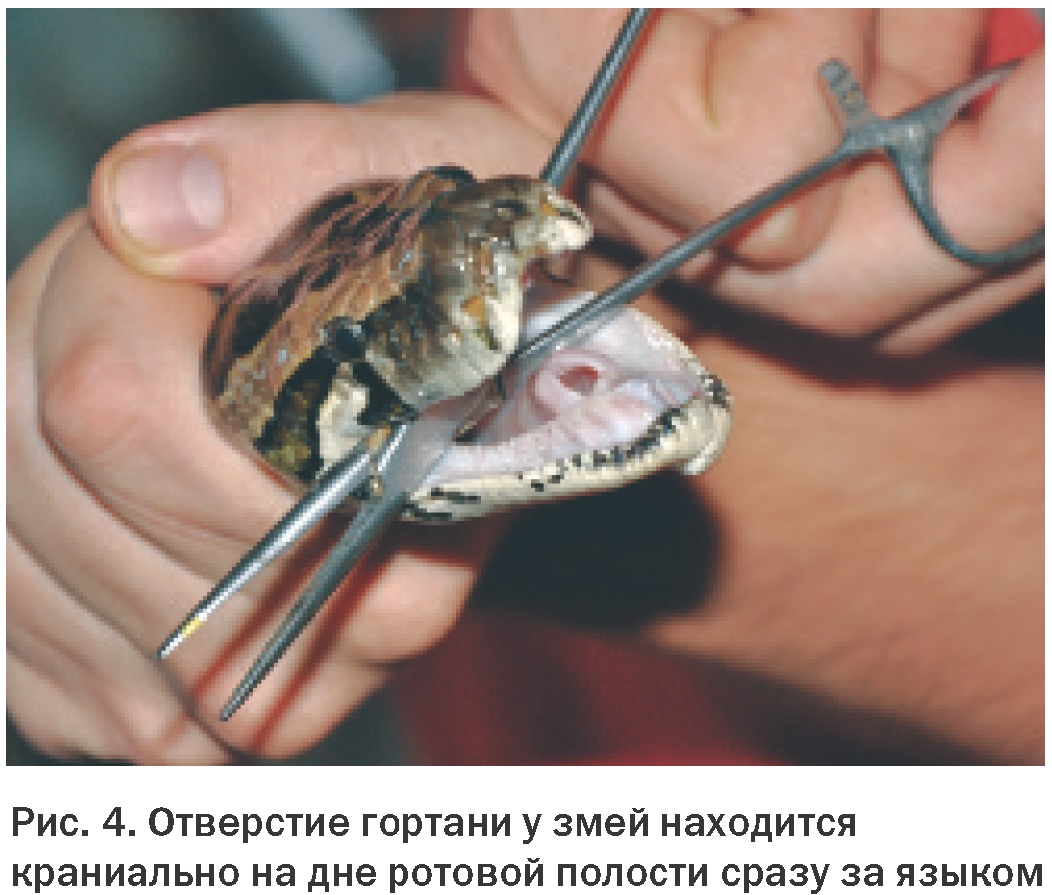
Отверстие гортани у змей расположено ближе к носу в ротовой полости, позволяя таким образом дышать во время заглатывания пищи (Рис. 4). Это также делает возможным прямой осмотр и интубацию трахеи у большинства змей без применения наркоза. Трахея с неполными С-образными кольцами входит в легкое на уровне нижней части сердца. У некоторых видов есть «трахейное легкое», расположенное на дорсальной стороне трахеи. Эта структура служит для продолжения газообмена в случае сдавливания легких проглоченной добычей.
У большинства змей левое легкое рудиментарно или отсутствует. У самых примитивных видов, таких как удавы, есть оба легких, но левое меньше. В легких большинства змей различают два отдела. Первая треть или половина легкого функционирует нормально, а задняя часть относительно мало пронизана кровеносными сосудами и похожа на воздушный мешок у птиц. Краниальная или дыхательная часть легкого обычно начинается на уровне 20% и заканчивается на уровне 40% от общей длины от носа до клоаки. Каудальная или мембранная часть легкого располагается далее и имеет разную длину у разных видов.
Дыхательный цикл змей включает активную и пассивную стадии. Выдох контролируется дорсолатеральным (поперечная мышца спины и верхняя внутренняя реберная мышца) и вентролатеральным (поперечная и косая мышцы живота) слоями мышц. Расслабление мышц, обеспечивающих выдох, является первым пассивным этапом вдоха. Подъём ребер в результате сокращения мышц, поднимающих ребро, и мышц, оттягивающих ребро, является активной частью вдоха. Последняя стадия дыхательного цикла, пассивный выдох, происходит в результате расслабления мышц вдоха и спадания легкого.
Крокодилы
Гортань у крокодилов открывается каудально за хорошо выраженным надгортанным клапаном. Вместе с подъязычной костью он может запирать ротовую полость, позволяя животному дышать при частичном погружении даже с открытым ртом. Полость внутренних органов у них разделена на две части мышечной перегородкой, аналогичной диафрагме. К ней примыкает краниальная часть печени. Каудально печень связана с тазовым поясом парой диафрагмальных мышц.
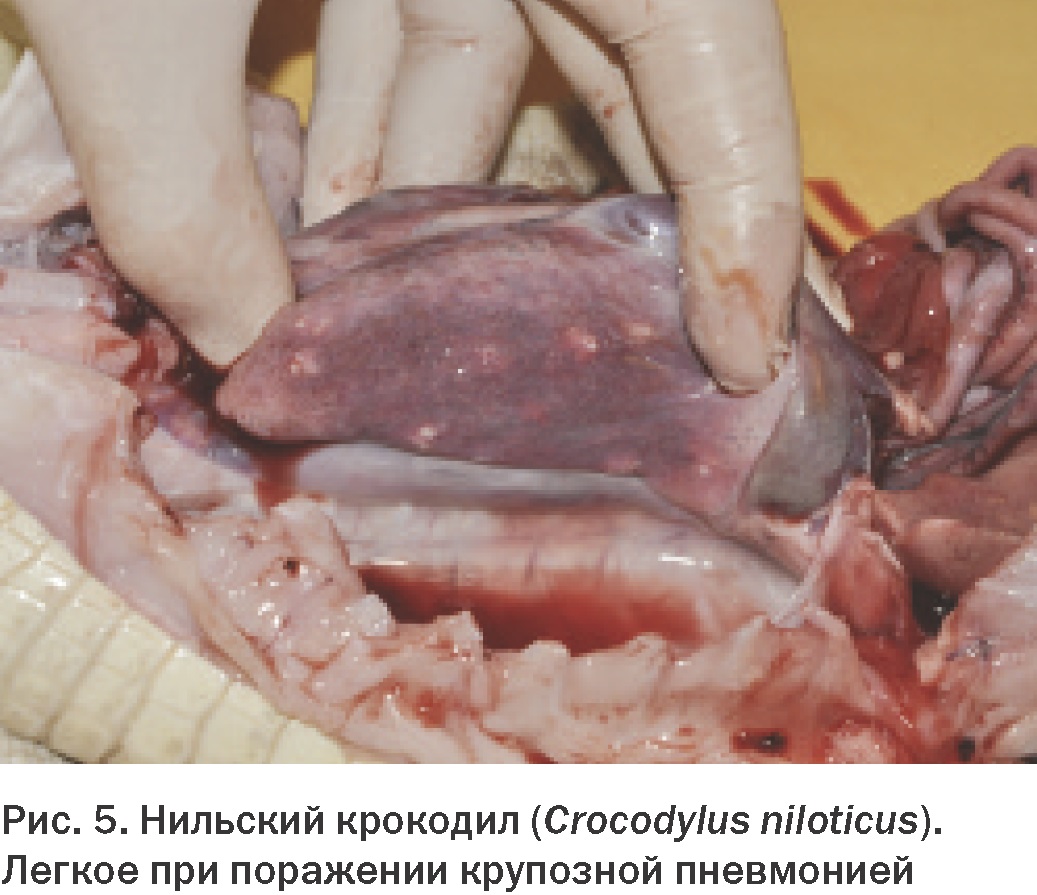
Сокращение этих мышц совместно с межреберными мышцами также направлено на вдох и часто называется «гепатическим поршнем». Активный выдох производится благодаря сжатию полости тела поперечными мышцами. Легкие крокодилов устроены сложнее, чем у других рептилий. Парабронхи делят их на ряд отдельных камер (Рис. 5).
Несмотря на огромные различия в общей анатомии дыхательной системы рептилий, ее микроскопическое строение довольно единообразно. Трахею выстилает многорядный, мерцательный, цилиндрический эпителий с различным количеством бокаловидных клеток. По окончанию крупных дыхательных путей большого диаметра мерцательный эпителий исчезает и заменяется чешуйчатым. Сетчатое альвеолярное легкое состоит из тонкостенных капилляров, проходящих между эпителиальными стенками фальвеол. Каудальное легкое без кровеносных сосудов выстлано простым чешуйчатым эпителием.
Физиология дыхания
Говоря о физиологии дыхания, следует рассмотреть три составляющие респираторного механизма: во-первых, движение респираторных газов из внешней среды к поверхности газообмена, во-вторых, перемещение кислорода и углекислого газа сквозь дыхательный эпителий и, в-третьих, циркуляцию крови, несущей кислород и углекислый газ, от легких к тканям и последующую диффузию газов на клеточном уровне.
Как было показано ранее, рептилии обладают уникальными системами ассимиляции и удаления респираторных газов. Хотя способы работы этих систем весьма вариабельны у разных видов рептилий, все остальные дыхательные процессы в целом одинаковы.
Важное клиническое значение имеет способность рептилий к анаэробному метаболизму. Особые буферные системы, присутствующие в кровеносной системе, позволяют управлять накоплением молочной кислоты и ионов водорода в периоды анаэробиоза. Из-за этой способности даже тяжелые заболевания дыхательной системы могут оставаться скрытыми, пока их прогрессия не превзойдет компенсаторные свойства организма.
Газообмен у рептилий не ограничивается легочными фальвеолами. Многие водные виды способны к газообмену через покровы. Такие черепахи, как мягкотелая черепаха (Apalone spp.), могут получать до 70% кислорода во время погружения через кожистый панцирь.
Щёчно-глоточные перепонки также являются поверхностями для газообмена у множества рептилий, а густо пронизанные кровеносными сосудами глоточные папиллы способны извлекать кислород, растворенный в воде. Мягкотелые черепахи могут получать достаточное количество кислорода через кожу и ротоглотку во время длительных погружений, например, во время спячки.
Многие виды ящериц также способны в некоторой степени к глоточному и кожному дыханию.
Как было упомянуто ранее, некоторые виды змей отличаются особым мешковидным выпячиванием открытых колец трахеи, которое служит для газообмена в том случае, если содержимое желудка мешает нормальной работе легкого. Дыхание пойкилотермных животных контролируется парциальным давлением кислорода, углекислого газа и температурой. При увеличении температуры потребность в кислороде также возрастает. Она обычно компенсируется не учащением дыхания, а увеличением дыхательного объема. Более того, гиперкапния (увеличение содержания CO2) также вызывает увеличение дыхательного объема, тогда как гипоксия приводит к учащению дыхания. Стимулом к учащенному дыханию у рептилий является низкая концентрация кислорода в крови.
При значительной легочной патологии способность пациента к увеличению дыхательного объема ухудшается из-за воспалительных процессов, клеточной инфильтрации и потери эластичности тканей.
Клинически важно, что условия с большим содержанием кислорода в воздухе ведут к подавлению собственных дыхательных движений у рептилий. Хотя отсутствие диафрагмы и слабый бронхоцилиарный транспорт ухудшает отход воспалительного экссудата из легких, длительная экспозиция в насыщенной кислородом среде еще более затруднит его в связи с подавлением вентиляции. Поэтому использование кислородных камер следует применять только в случаях подозрения на гипоксемию. Относительная неэффективность легких в сочетании со способностью рептилий переключаться на анаэробный тип обмена веществ играют важную роль при патологиях дыхательной системы.
Серьезные респираторные заболевания могут вызывать увеличение сопротивления легочных кровеносных сосудов. Так как эти изменения сходны с таковыми при задержке дыхания или погружении, они ведут к изменениям в работе сердца. При нормальном дыхании сердце рептилий, кроме крокодилов, направляет кровь, наиболее бедную кислородом, в легочную артерию. Увеличение сопротивления легочных сосудов приводит к отводу крови от легких в системный круг кровообращения. Если отвод возникает в результате патологии, перенос кислорода к периферическим тканям может быть затруднен. При продолжительном анаэробиозе в результате перегрузки буферной системы крови развивается ацидоз, представляющий угрозу для жизни. ■
Литература
1. Jackson CG Jr: Cardiovascular system. In Harless M, Morlock H, editors: Turtles: perspectives and research, Melbourne, Fla, 1979, Krieger Publishing.
2. White FN: Circulation. In Gans C, editor: Biology of the reptilia, vol 5, New York, 1976, Academic Press.
3. Williams DL: Cardiovascular system. In Beynon PH, Law-ton MPC, Cooper JE, editors: Manual of reptiles, Cheltenham, United Kingdom, 1992, British Small Animal Veterinary Association.
4. Smith JA, McGuire NC, Mitchell MA: Cardiopulmonary physiology and anesthesia in crocodilians, Proc Assoc Reptil Amphib Vet 17-21, 1998.
5. Holz PH: The reptilian renal portal system — a review, Bull Assn Reptil Amphib Vet 9(1):4-9, 1999.
6. Davies PMC: Anatomy and physiology. In Cooper JE, Jackson OF, editors: Diseases of the Reptilia, vol 1, New York, 1981, Academic Press.
7. Anderson JB, Rourke BC, Caiozzo VJ, Bennett AF, Hicks JW: Postprandial cardiac hypertrophy in pythons, Nature (434): 37-38, 2005.

Назад в раздел














