

Грибы рода malassezia в заболеваниях животных: биология, экология, патогенез, предрасполагающие факторы

Еще фото
Автор (ы): Р.С. Овчинников
Организация(и): Федеральное государственное бюджетное учреждение «Всероссийский государственный Центр качества и стандартизации лекарственных средств для животных и кормов» (ФГБУ «ВГНКИ»)
Журнал: №1 - 2013
Введение
В последние десятилетия как в ветеринарии, так и в медицине возросло число микозов, развивающихся на фоне снижения резистентности макроорганизма, для обозначения которых был введен термин «оппортунистические микозы». Учитывая разнообразие негативных факторов, оказывающих постоянное воздействие на организм животных в современных условиях и ослабляющих их иммунный и физиологический статус, тенденция к распространению оппортунистических микозов представляется закономерной.
Оппортунистические микозы животных, вызываемые дрожжевыми грибами рода Malassezia, начиная с 1990-х гг., стали одной из наиболее насущных проблем в ветеринарной микологии [1-3]. Malassezia-инфекции имеют повсеместное распространение, им подвержены многие виды домашних животных, в то время как существующие методы диагностики и терапии этих микозов недостаточно эффективны [4].
Вплоть до середины 2000-х гг. на территории РФ распространенность и этиологическая структура Malassezia-инфекций животных практически не изучалась; они очень слабо освещены в русскоязычной литературе. Информированность практикующих ветеринарных врачей и сотрудников лабораторий о данных заболеваниях находится в целом на низком уровне, в силу чего не обеспечивается должный уровень диагностики и терапии.
В данной обзорной статье обобщены современные научные данные, касающиеся роли грибов рода Malassezia в заболеваниях животных. В первой части рассмотрены вопросы биологии и экологии возбудителя, механизмы патогенеза, значение предрасполагающих факторов. Продолжение статьи будет посвящено клиническим аспектам, диагностике и терапии малассезиозов.
История открытия, классификация, номенклатура
Роль грибов рода Malassezia в кожных заболеваниях человека была установлена в середине XIX века. Первое описание дрожжевых грибов, ассоциированных с пестрым лишаем человека (Tinea versicolor), принадлежит E. Eichstedt. В 1853 г. Robin описал округлые дрожжевые клетки, обнаруженные им в кожных чешуйках, и присвоил им видовое название Microsporon furfur. Ветеринарное значение этих микроорганизмов было выявлено значительно позже.
В 1925 г. Weidman [5] в кожных чешуйках от индийского носорога с клиникой дерматита обнаружил дрожжеподобные клетки бутылевидной формы. Weidman выделил культуру и отнёс микроорганизм к роду Pityrosporum, т.к. по описанию он напоминал вид P. оvale, выделенный от людей. Однако выделенная культура имела ряд отличий, что дало Weidman основание выделить ее в отдельный новый вид P. pachydermatis.
В 1955 г. Gustafson [6] впервые выделил дрожжевую культуру с бутылевидными клетками от собаки с наружным отитом и описал ее как новый вид рода Pityrosporum – P. canis. Он впервые предположил, что этот микроорганизм играет роль в патогенезе отита. После этого длительное время считалось, что ветеринарное значение Malassezia spp. только этим и ограничивается.
Гораздо позже R. Dufaut (1983) установил этиологическую значимость этих грибов при дерматитах собак, в т.ч. генерализованного характера [7]. С середины 80-х гг. вопрос Malassezia-ассоциированных заболеваний животных попал в фокус внимания как врачей-практиков, так и исследователей, стал целенаправленно изучаться и активно обсуждаться в научной литературе [8-11].
Прежнее название рода Malassezia – Pityrosporum – было предложено Sabouraud в 1904 г., а в 1952 г. былf официально принята таксономическая классификация рода Pityrosporum, предложенная J. Lodder. C 1986 г. официально утверждено новое название рода – Malassezia Baillon. По современной классификации род Malassezia Baillon относится к отделу Basidiomycota, подотделу Ustilagomycotina, классу Exobasidiomycetes, порядку Malasseziales, семейству Malasseziaсeae [12].
К настоящему времени описано 14 видов этого рода: M. furfur, M. pachydermatis, M. sympodialis, M. globosa, M. obtusa, M. restricta, M. slooffiae, M. dermatis, M. japonica, M. nana, M. yamatoensis, M. caprae, M. equina, M. cuniculi, причем последние 7 видов были открыты уже в 2000-х годах. Все виды данного рода выделены от животных и человека. Основное ветеринарное значение имеет вид M. pachydermatis.
Морфология, физиология, факторы патогенности
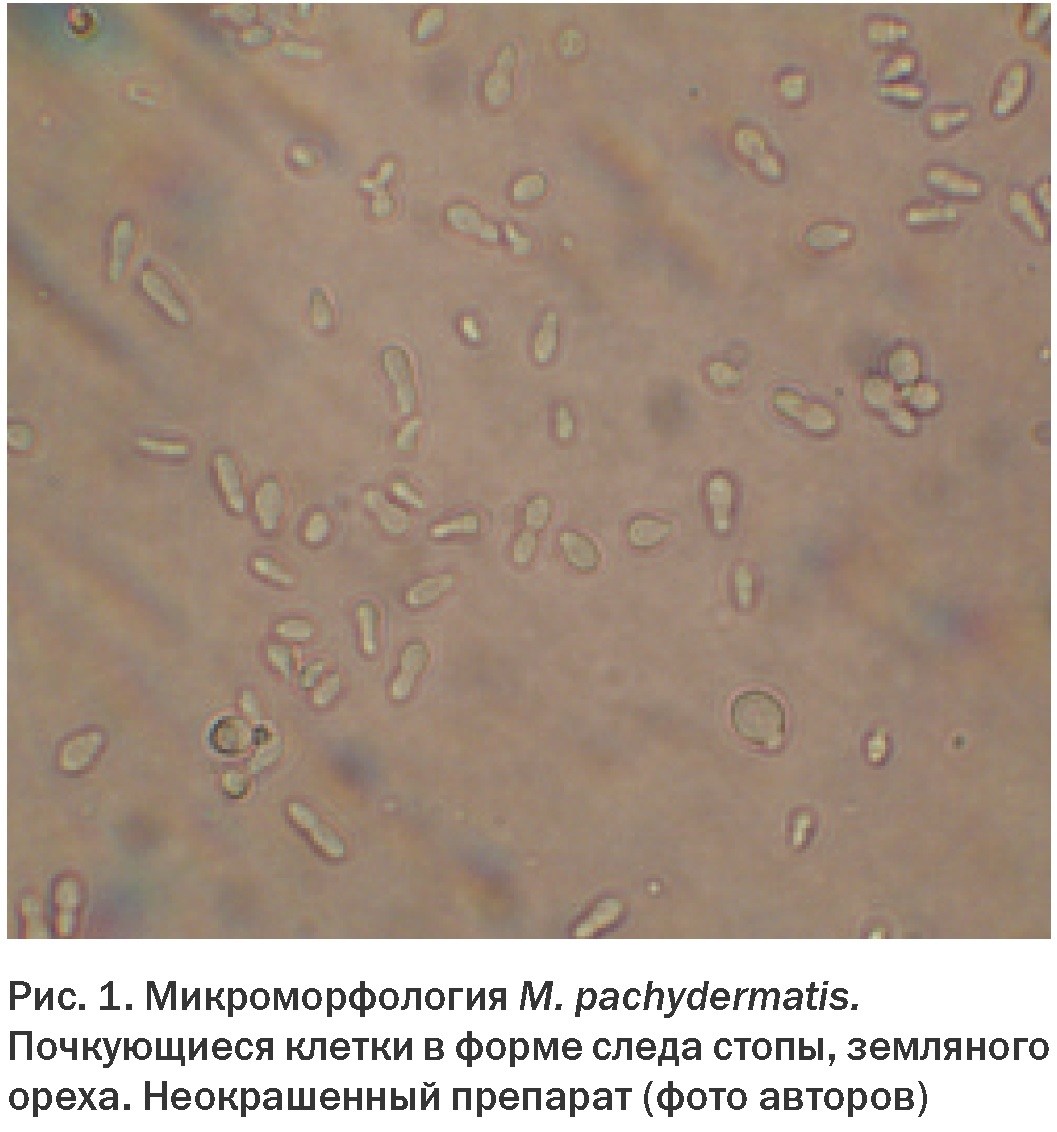
Для грибов рода Malassezia характерны определенные морфологические и физиологические особенности. У M. pachydermatis клетки овальной, эллиптической, реже округлой формы, размер клеток 2-3 х 3-5 мкм (Рис. 1). Для других видов характерна округлая, лимоновидная форма клеток. В дрожжевой стадии тип почкования монополярный, перкуррентный – отделившаяся почка оставляет на материнской клетке шрам в виде воротничка, из которого появляются последующие почки. В силу особенностей почкования клетки приобретают характерную грушевидную форму или форму «отпечатка стопы» или «земляного ореха». По Граму окрашиваются положительно (Рис. 2).
Грибы рода Malassezia могут существовать как в дрожжевой, так и в мицелиальной форме (однако у вида M. pachydermatis мицелиальная форма неизвестна). В культуре, как правило, доминирует дрожжевая форма, в то время как мицелиальная ассоциирована с инвазивной фазой роста гриба.
Эти микроорганизмы характеризуются толстой многослойной клеточной стенкой (толщиной до 0,25 мкм) со складчатой внутренней поверхностью. Толстая клеточная стенка обеспечивает устойчивость дрожжей к воздействию защитных факторов макроорганизма, благодаря чему эти грибы способны выживать и персистировать на кожном покрове теплокровных животных. Внешняя поверхность клеточной стенки M. pachydermatis покрыта тонким ламеллярным слоем, содержащим липиды, который играет роль в адгезии дрожжей к клеткам кожи [13] (Рис. 3). Примечательно, что адгезия к корнеоцитам M. pachydermatis происходит в 10 раз активнее, чем у Saccharomyces cerevisae (пекарские дрожжи) [14].
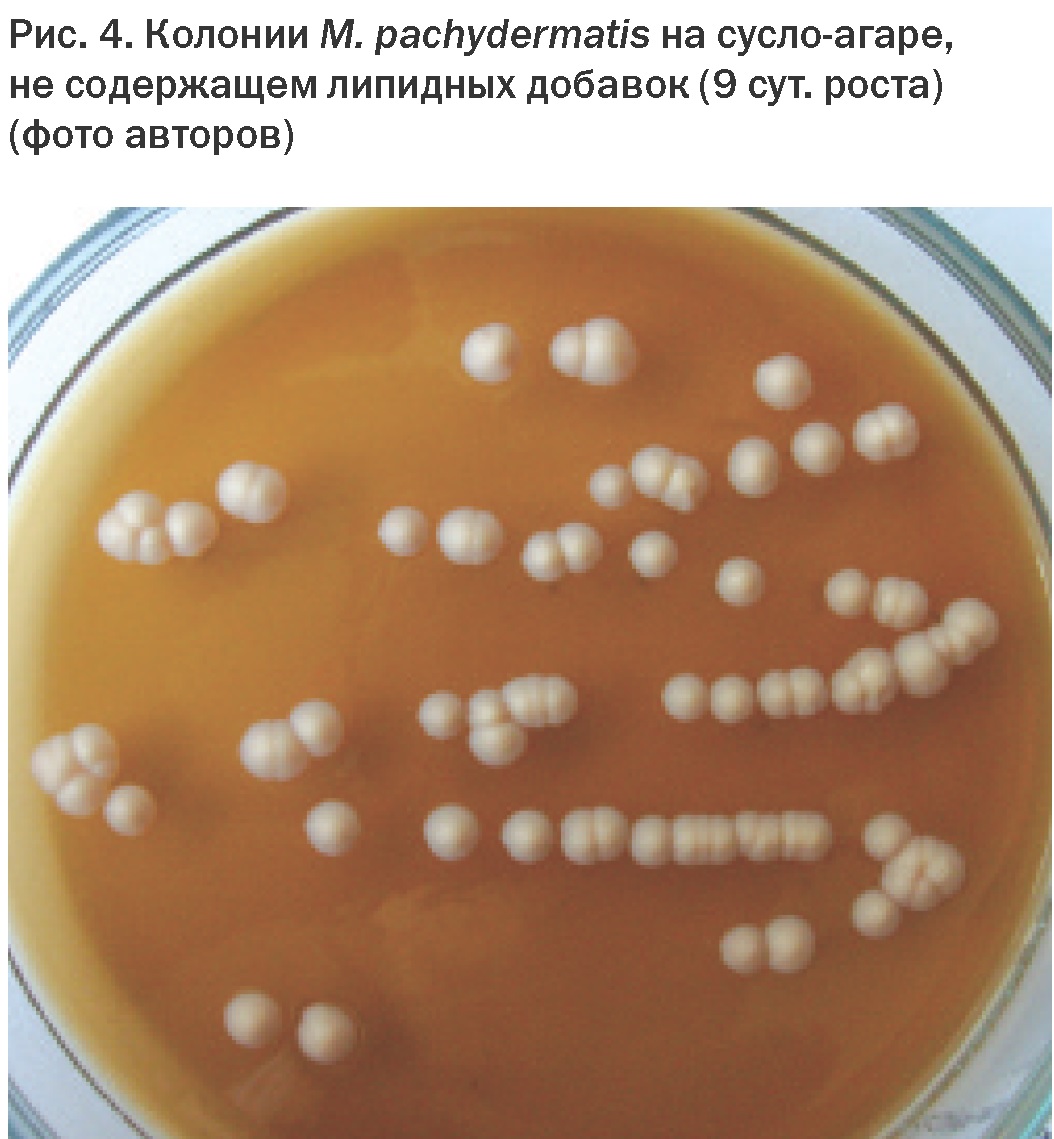
Длительное время считалось, что дрожжи рода Malassezia вообще не могут быть выделены в культуру, т.к. была неизвестна их липид-зависимость. В 1913 г. Castellani и Chalmes выделили культуру гриба и впервые охарактеризовали его ростовые свойства. Липофильность – уникальная особенность грибов этого рода. Потребность этих микроорганизмов в «жироподобных веществах» была изучена R. Benham [15]. На средах, не содержащих липидов, способен расти лишь один вид – M. pachydermatis, однако в присутствии липидов его рост существенно стимулируется [16] (Рис. 4).
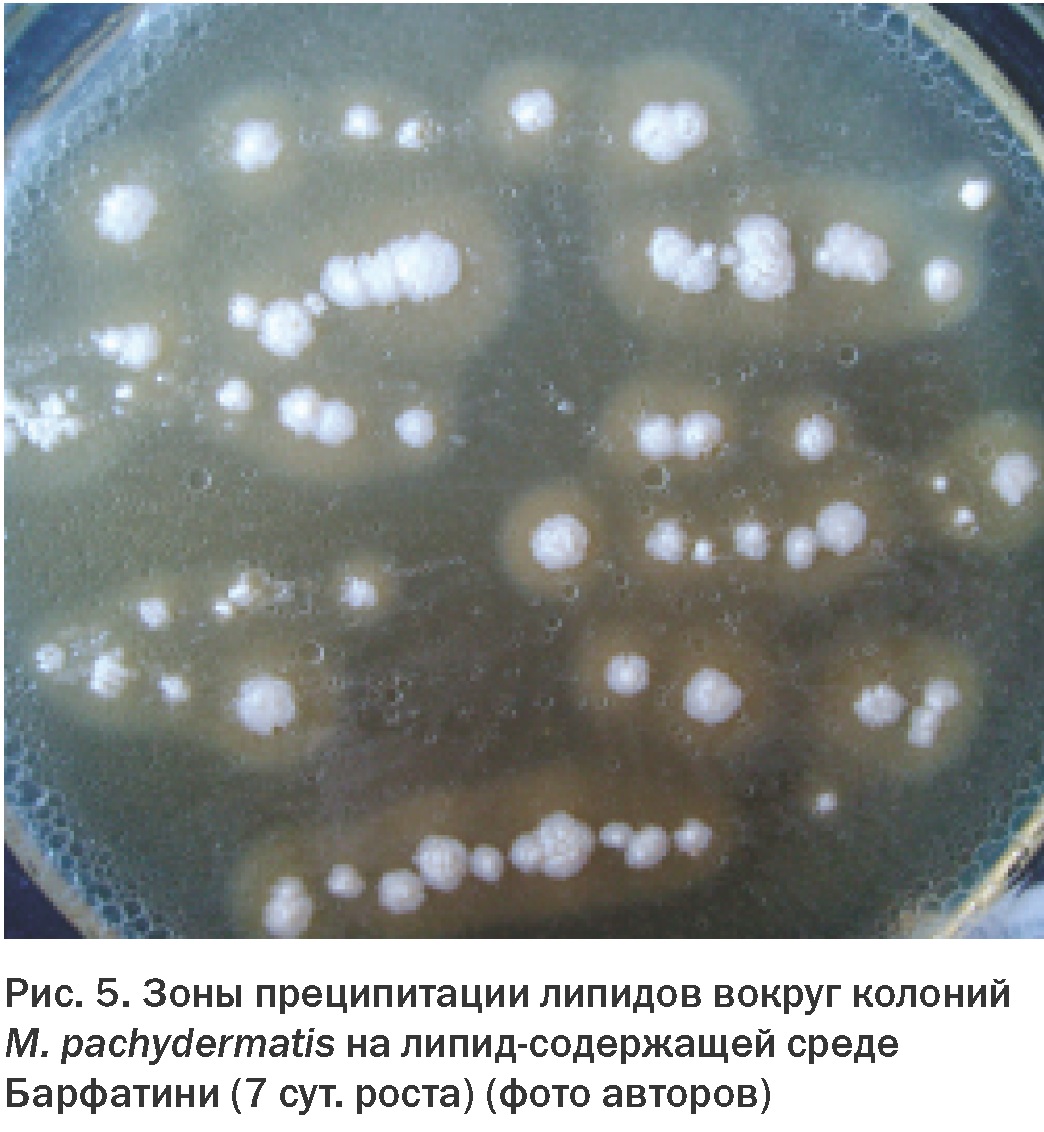
Позже было установлено, что для роста грибов рода Malassezia необходимо наличие в питательной среде жирных кислот с длиной цепи С12-С24, которые грибы рода Malassezia не способны синтезировать самостоятельно. Эти компоненты встречаются в натуральных маслах – оливковом, кукурузном, соевом, подсолнечном [17]. Исходя из этого, для выделения липофильных дрожжей используют специальные липидсодержащие питательные среды. На таких средах можно наблюдать образование зон преципитации липидов вокруг колоний малассезий, что может служить дополнительным дифференциально-диагностическим признаком (Рис. 5).
Липофильность обуславливается наличием липолитических ферментов, способных гидролизовать липиды кожного секрета до свободных жирных кислот. В свою очередь, свободные жирные кислоты ингибируют рост других микроорганизмов, повышая конкурентоспособность дрожжей. C. Cafarcia et al. [18] полагают, что фосфолипазы – ферменты, разрушающие эфирные связи в глицерофосфолипидах – играют существенную роль в патогенности Malassezia spp. Фосфолипазная активность была обнаружена у 93,9% изолятов M. pachydermatis, выделенных от собак с клиникой Malassezia-дерматита, в то время как изоляты с кожи здоровых животных проявили активность только в 10,6% случаев.
Помимо липаз, M. pachydermatis синтезирует протеиназу, сульфатазу, гиалуронидазу, которые также играют роль в пенетрации покровных тканей организма-хозяина [19]. Chowdhary A. et al. [20] впервые сообщили о кератинолитических свойствах Malassezia furfur на основании зарегистрированного ими случая онихомикоза, обусловленного данным видом гриба.
Экспериментально продемонстрирована способность Malassezia spp. проникать в роговой слой, осуществляя его деструкцию, что говорит о способности к инфекционной инвазии. Однако проникновение дрожжевых клеток, по данным авторов, ограничивается лишь верхними слоями эпидермиса [21].
Грибы рода Malassezia вызывают различные иммунные реакции в макроорганизме, причем иммуногенные свойства во многом обусловлены липидным слоем этих грибов. Они взаимодействуют с различными иммунокомпетентными клетками, включая антиген-презентующие дендритные клетки, макрофаги, эозинофилы, нейтрофилы, стимулируя синтез цитокинов и хемокинов различных функциональных групп. Установлено, что малассезии способны как стимулировать, так и ингибировать синтез провоспалительных цитокинов (IL-1, IL-6, TNF- ) [22]. Вследствие этого воспалительная реакция при малассезиозах может варьировать от практически незаметной до ярко выраженной.
Виды рода Malassezia способны связываться с лектинами С-типа (разнородная группа протеиновых рецепторов, которые способны распознавать углеводные структуры микроорганизмов) и вызывать клеточный иммунный ответ.
Малассезии индуцируют синтез цитокинов как через Toll-подобный рецептор 2 (TLR2/MyD88-механизм), так и через другие, пока точно не выявленные механизмы [12].
Как отмечает H. Ashbee [23], грибам рода Malassezia присуща парадоксальная способность как стимулировать, так и ингибировать иммунные механизмы организма-хозяина, что позволяет этим грибам балансировать между комменсализмом и паразитизмом.
Экология грибов рода Malassezia
В отличие от других представителей клинически значимых дрожжевых грибов, малассезии не являются убиквитарными организмами и практически не выделяются из объектов внешней среды, за редкими исключениями [24].
Грибы рода Malassezia – представители нормальной микробиоты кожи и слизистых большинства теплокровных животных и человека. Успешной пролиферации этих грибов на коже благоприятствуют участки тела с повышенной температурой, влажностью, повышенным содержанием СО2, наличием липид-содержащего кожного секрета, являющегося для дрожжей источником питательных веществ. Так, у собак грибы рода Malassezia чаще всего выделяли из области губ (80% животных), межпальцевых складок (60%), наружного слухового прохода (32,5%), со слизистых оболочек (52%) [25]. Кроме того, этим грибам свойственны высокая осмотолерантность, галофильность, значение рН оптимума 5,5, что обеспечивает успешное существование на кожном покрове [26].
В норме взаимоотношения грибов рода Malassezia и организма-хозяина носят характер симбиоза. Обитая на кожном покрове здоровых животных, дрожжи потребляют жироподобные вещества, содержащиеся в кожных секретах, и, в свою очередь, синтезируют антибактериальные и антифунгальные вещества, препятствующие развитию на коже других патогенных микроорганизмов. Так, продукты метаболизма M. furfur обладают антифунгальной активностью в отношении патогенных грибов-дерматофитов [27].
Колонизация кожного покрова происходит в раннем послеродовом периоде. У собак дрожжи обнаруживают уже в 3-дневном возрасте в области губ, ногтевого ложа, наружного слухового канала. Предполагают, что передача дрожжей происходит от матери к потомству посредством вылизывания, груминга, или со слизистых оболочек вагины. Вероятно, поверхность слизистых оболочек является естественным резервуаром Malassezia, откуда дрожжи впоследствии распространяются на другие участки тела [28].
Malassezia-носительство широко распространено у клинически здоровых животных. Встречаемость M. pachydermatis в слуховом канале здоровых собак составляет, по разным данным, от 2 до 49% и более. У 40% собак этот вид обнаруживали в анусе, прямой кишке, перианальных железах, вагине. На шерстном покрове встречаемость Malassezia spp. варьирует в зависимости от локуса. Грибы часто обнаруживают в межпальцевых пространствах, на волосистой коже нижней губы, в отличие от подмышек и паха [14]. Жизнеспособные клетки грибов рода Malassezia были обнаружены в фекалиях собак [29].
По данным Guillot et Bond [14], степень к грибной колонизации зависит и от породы – так, у бассет-хаундов Malassezia-носительство на коже и слизистых встречается гораздо чаще, нежели у других пород собак. В то же время С. Reme [30] сообщает, что наличие и численность популяции малассезий не коррелировали с такими факторами, как порода, тип ушной раковины (стоячая или висячая), пол, возраст. Этот автор обнаружил Malassezia-носительство в слуховом канале 71% здоровых собак.
Среди кошек Malassezia-носительство также широко распространено. S. Colombo et al. [31] выявили кожное миконосительство у 61% здоровых кошек, а у кошек породы девонрекс – в 100% случаев. R. Bond et al. [32] диагностировали кожное Malassezia-носительство у всех обследованных кошек. C. Cafarcia et al. [33] выделяли грибы рода Malassezia от 28% обследованных собак и 40% кошек. J. Guillot et al. [34] изолировали M. pachydermatis из слухового прохода здоровых кошек и собак в 20% и 42% случаев соответственно.
Кожный покров сельскохозяйственных животных также часто бывает колонизирован грибами рода Malassezia. M. Crespo et al. [35] установили, что среди лошадей Malassezia-носительство имеет место у 60% животных, среди крупного рогатого скота – у 58%, среди овец – у 28%, среди коз – у 44%. M. Garau et al. [36] выделяли виды M. sympodialis и M. slooffiae из слухового канала здоровых свиней в 73% случаев.
Имеются сообщения о выделении грибов рода Malassezia с кожного покрова хорьков, приматов, бурых медведей, вомбатов, барсуков, индийских слонов, носорогов, кабанов, лисиц, дикобразов, койотов, ластоногих, попугаев [14, 37].
Патогенетическое значение грибов рода Malassezia
Болезнетворные свойства грибов рода Malassezia и их клиническая роль в заболеваниях животных до сих пор трактуются неоднозначно. Нет единого мнения, могут ли Malassezia-инфекции рассматриваться как самостоятельное заболевание, или же они являются лишь отягощающим фактором на фоне других патологий.
В медицинской микологии убедительно доказана этиологическая роль грибов рода Malassezia при пестром лишае. Также эти грибы участвуют в патогенезе себорейного дерматита, атопического дерматита, фолликулитов, псориаза и других кожных заболеваний [17, 38, 39, 40]. Известны случаи фунгемий и других системных микозов, вызванных Malassezia spp. [41].
Интенсивные исследования последних десятилетий внесли существенный вклад в представление о патогенезе Malassezia-инфекций. В настоящее время большинство исследователей разделяет мнение, что патогенность грибов рода Malassezia носит «оппортунистический» характер, т.е. гриб способен проявлять патогенные свойства только на фоне определенных предрасполагающих факторов [42, 43].
Развитие патологического процесса при малассезиозах связано с многократным увеличением популяции микроорганизмов в очаге поражения. G. Fraser [44] выделял M. pachydermatis от здоровых собак и собак с отитом примерно в одинаковых пропорциях, однако он заметил, что у больных собак количество дрожжевых клеток значительно больше. Дальнейшие исследования подтвердили эту закономерность.
По данным R. Bond [45], популяция дрожжей у больных животных повышается в 100-10000 раз. Причем отмечают рост численности дрожжей не только на поверхности кожи, но и на слизистых носовой полости, вульве и препуции, т.е. предрасполагающие к этому факторы носят не местный, а системный характер.
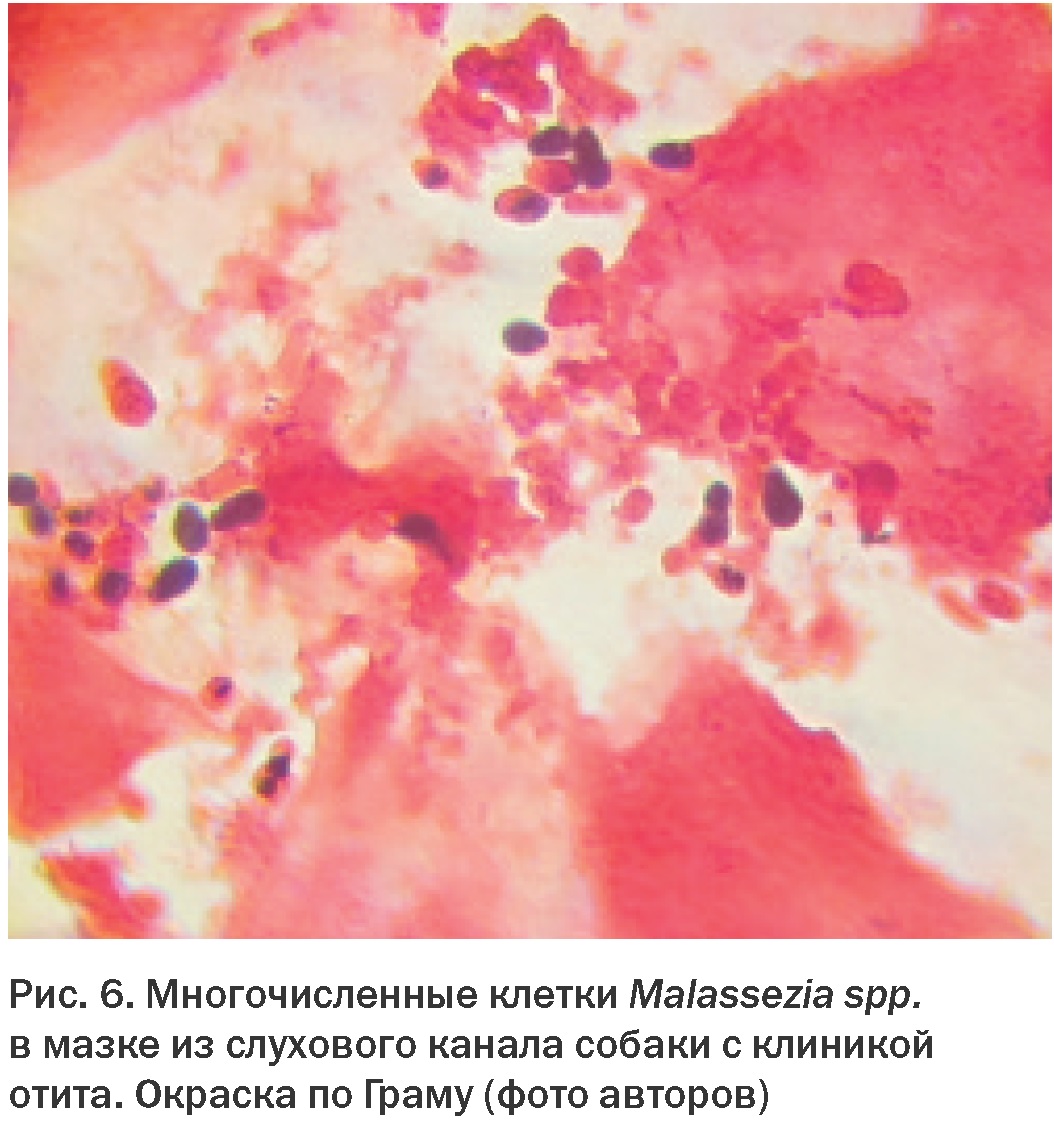
По данным наших собственных исследований, плотность популяции Malassezia spp. в слуховом канале здоровых собак составляет в среднем 7,0 КОЕ в пробе, при этом дрожжи были выделены у 29,6% обследованных животных. В то же время у собак с клиникой отита плотность популяции составляла в среднем 216 КОЕ при частоте выделения 88,2% [46]. Также большое количество дрожжевых клеток обнаруживают и при цитологическом исследовании материала (Рис. 6).
По современным представлениям, в запуске патологического процесса ключевую роль играет коммуникация микроорганизмов внутри популяции, осуществляющаяся посредством сигнальных молекул. Предполагается, что когда популяция достигает определенной численности, в ней возникает «чувство кворума», что служит стартовым сигналом для активации факторов патогенности и, как следствие, ведет к развитию инфекционного процесса [47].
С механизмом внутрипопуляционной коммуникации микроорганизмов связано такое явление, как образование биопленок – надорганизменных структур, обеспечивающих защитные и трофические функции. Известно, что способность к образованию биопленок является одним из факторов патогенности дрожжевых грибов рода Candida. В 2007 г. F. Cannizzo et al. установили, что грибы рода Malassezia также способны образовывать биопленки на поверхности различных субстратов [48].
По данным J. Faergemann [49], патогенность грибов рода Malassezia также обуславливается механизмом филаментации – переходом дрожжевых клеток в мицелиальную форму. Этот же механизм лежит в основе патогенности дрожжевых грибов рода Candida. Однако известно, что вид M. pachydermatis существует только в дрожжевой форме.
Установлено, что в патогенезе заболевания играют роль симбионтные отношения дрожжей и обитающих на коже бактерий (в частности, стафилококков). Стафилококки также продуцируют липазу, что нарушает секреторную функцию кожи и создает благоприятные условия для роста обоих организмов, в то время как для других конкурентных микроорганизмов такие условия неблагоприятны. Помимо стафилококков, в патогенезе заболевания могут играть роль другие виды бактерий и микроскопических грибов. A. Kumar et al. от собак, больных отитом, наряду с M. pachydermatis (83 изолята) выделяли бактерии – Staphylococcus spp. (70), Pseudomonas spp. (27), Proteus spp. (13), Streptococcus spp. (7), а также грибы – Candida spp. (13) и Aspergillus spp. (9) [50]. F. Breitwieser [51] сообщает, что в слуховом проходе больных отитом собак в 31,8% случаев M. pachydermatis был ассоциирован с Staphylococcus spp., в 22,9% – с Pseudomonas aeruginosa, в 8% – с Streptococcus spp.
Предрасполагающие факторы при Malassezia-инфекциях
Факторы, способствующие возникновению Malassezia-инфекций, могут быть самой разнообразной природы. Как отмечают P. Ginel et al. [52], кожа, особенно в слуховом канале, представляет собой хрупкую многокомпонентную систему, и даже небольшие изменения в кожном микроклимате способны нарушить тонкий баланс в секреторном механизме и составе микрофлоры, что, в свою очередь, может привести к возникновению оппортунистических инфекций.
R. Bond et al. [53] экспериментальным путем продемонстрировали, что в возникновении малассезиозов важную роль играет микроклимат кожи (повышенная температура, влажность, концентрация СО2). При экспериментальном создании соответствующих условий (локальной окклюзии), авторы наблюдали «самопроизвольное» возникновение Malassezia-дерматита у собак за счет резкого увеличения резидентной кожной популяции M. pachydermatis.
Во многих случаях малассезиозы возникают на фоне первичных заболеваний и нарушений, таких как:
- иммунные дисфункции (кожная гиперчувствительность, атопический дерматит, пищевые, контактные аллергии);
- пиодерма;
- эндокринные нарушения (особенно гипотироидизм; гиперадренокортицизм, диабет);
- нарушения метаболизма (цинкзависимый дерматоз, поверхностный некролитический дерматит);
- нарушения кератинизации (эпидермальная дисплазия, идиопатическая себорея);
- лечение глюкокортикоидами или антибиотиками [4, 28].
По ряду данных, основной патологией, на фоне которой может развиться Malassezia-инфекция, является атопический дерматит – многофакторное заболевание, сопровождающееся аномальными реакциями иммунитета [54]. T. Nuttall et al. [55] установили, что у больных атопическим дерматитом собак уровень Malassezia-специфических иммуноглобулинов IgG и IgM существенно выше, нежели у здоровых животных. Малассезии проявляют аллергизирующие свойства – 2/3 собак с «себорейным дерматитом» проявляют реакцию гиперчувствительности немедленного типа при введении экстрактов из Malassezia spp. [56].
По данным R. Bond et al. [57], у 27 из 40 собак с Malassezia-инфекцией диагностировали также сопутствующие (конкурирующие) заболевания, в т.ч. ато-пический дерматит (15 случаев), нарушения кератинизации, эндокринопатии.
В наших собственных исследованиях наиболее распространенными сопутствующими заболеваниями были пищевые аллергии (63%), атопический дерматит (53%), эндокринные патологии (гипотироидизм, гиперадренокортицизм) (42%) [46].
P. Sierra et al. [58] установили, что грибы рода Malassezia существенно чаще выделяются с кожного покрова кошек, инфицированных вирусами FIV, FeLV, нежели от здоровых животных.
Малассезиозы могут быть ассоциированы с паразитарными заболеваниями. Z. Radi [59] сообщает о вспышке генерализованного инфекционного дерматита среди кроликов, вызванного ассоциацией грибов рода Malassezia и клещей рода Sarcoptes.
Предполагается роль малассезий в развитии некоторых типов опухолей, в частности, базальной клеточной карциномы. У кошек и собак такие опухоли образуются в области шеи и головы – местах, наиболее часто колонизируемых липофильными дрожжами. К ним более предрасположены породы собак с обильным шерстным покровом и висячими ушами (сенбернары, скотч-терьеры), чьи анатомические особенности благоприятствуют обильному росту малассезий [12].
Во многих исследованиях установлена корреляция заболеваемости малассезиозами и породной принадлежности животных. Среди собак Malassezia-отиты чаще регистрируются у пород с длинными висячими ушными раковинами: бассет-хаунд, коккер-спаниель, пудель, белый терьер, сеттер, далматин, бигль [57]. Это связывают с анатомическими особенностями наружного уха. Висячие ушные раковины затрудняют вентиляцию слухового прохода. Этому же способствует рост шерсти в слуховом проходе, наблюдаемый у некоторых пород (коккер-спаниели, бассеты). Задержке серы в слуховом проходе способствует стеноз ушного канала, что характерно для шарпеев и ротвейлеров. К затруднению нормального отделения ушной серы приводит и разрастание хрящевой части ушного канала, что встречается, в частности, у американских коккер-спаниелей [60].
G. Fernandez et al. [61] отмечают высокую распространенность Malassezia-отитов среди пуделей (30,1%), коккер-спаниелей (26,4%), терьеров (16,7%), германских шепердов (9,4%). Авторы связывают эту закономерность с наличием волосяного покрова в ушной раковине у пуделей и терьеров и гиперактивностью апорциновых желез ушного канала у коккер-спаниелей и шепердов. Однако по данным C. Reme [30], тип и строение ушной раковины не влияют на предрасположенность к Malassezia-инфекциям. По мнению автора, гораздо более важным фактором является температура в наружном ушном канале, а она, в свою очередь, зависит от веса животного. Так, у некрупных пород температура в слуховом канале относительно невысокая, что, очевидно, связано с более быстрой потерей тепла через кожу ушной раковины.
В наших собственных исследованиях установлена относительно высокая встречаемость малассезиозов у таких пород, как французский и английский бульдоги, мопс, шарпей, чау-чау. Все они относятся к брахицефалам – породам с укороченной мордой, т.е. эта анатомическая особенность строения черепа в определенной мере коррелирует с предрасположенностью животных к Malassezia-инфекциям [46].
В целом в большинстве исследований прослеживается взаимосвязь между породой и предрасположенностью собак к Malassezia-инфекциям. Однако вопрос, какие именно особенности породы играют роль предрасполагающего фактора, требует дальнейшего изучения. Также надо принимать во внимание распространенность и популярность тех или иных пород в конкретной местности.
У кошек к Malassezia-инфекциям предрасположены породы с абнормальным шерстным покровом – сфинкс, девон-рекс [62]. Среди пород крупного рогатого скота интенсивная колонизация слухового прохода грибами рода Malassezia чаще обнаруживается у зебувидных пород, для которых характерны висячие ушные раковины [63].
Экзогенные факторы, в частности, климатические условия, также могут способствовать заболеваемости Malassezia-инфекциями. Заболевание чаще регистрируется в летние месяцы, в периоды повышенной температуры и влажности воздуха [4, 50, 63]. По данным M. Saridomichelakis et al. [64], низкая (2,1%) распространенность этих заболеваний среди собак в Греции обусловлена сухим климатом этой страны. С другой стороны, в Венесуэле, стране с тропическим климатом, распространенность Malassezia-отитов среди собак составила 69,8% [61].
Таким образом, механизм патогенеза Malassezia-инфекций обусловлен, с одной стороны, эндогенными и экзогенными предрасполагающими факторами, а с другой – патогенным потенциалом дрожжевых грибов (особенностями строения грибной клетки, способностью к адгезии, грибными гидролитическими ферментами, механизмом филаментации, взаимодействием с другими патогенными микроорганизмами). Однако механизмы активации факторов патогенности и иммунного ответа при Malassezia-инфекциях требуют дальнейшего изучения.
Литература
1. Dorogi J. Pathological and clinical aspects of the diseases caused by Malassezia species // Acta Microbiol. Immunol. Hung. - 49 (2-3), 2002. - P. 363-369.
2. Weiss R., Raabe P., Mayser P. Yeasts of the genus Malassezia: taxonomic classification and significance in (veterinary and) clinical medicine // Mycoses. - 43 (Suppl. 1), 2000. - Р. 69-72.
3. Batra R., Boekhout T., Gueho E., Cabanes F., Dawson T. Malassezia Baillon, emerging clinical yeasts // FEMS Yeast Res. - 5(12), 2005. - Р. 1101-1113.
4. Patterson A., Frank L. How to diagnose and treat Malassezia dermatitis in dogs // Vet. Med. - August 1, 2002. - Р. 612-622.
5. Weidman F. Exfoliative dermatitis in the Indian rhinoceros (Rhinoceros inicornis), with dscription of a new species: Pityrosporum pachydermatis // In: Fox H., ed. Rep. Lab. Mus. Comp. Zoo Soc. Philadelphia. 1925. - P. 36-43.
6. Gustafson B. Otitis externa in the dog. A bacteriological and experimental study // Thesis, Royal Veterinary College of Sweden. Stokholm, 1955. - P. 117.
7. Dufaut R. Pityrosporum canis as the cause of canine chronic dermatitis // Vet. Med. Small Anim. Clin. - 78, 1983. - P. 1055-1057.
8. Dufaut R. Presence de Malassezia pachydermatis (syn. Pityrosporum canis) sur les poils et les plumes des animaux domestiques // Bull. Soc. Fr. Mycol. Med. - 14, 1985. – P 19-22.
9. Bliss E. Tinea versicolor dermatomycosis in the goat // J. am. Vet. Med. Assoc. - 184, 1984. - Р. 1512-1513.
10. Larsson C., Gandra C., Larsson M. et al. Dermatitis in dogs caused by Malassezia (Pityrosporum) pachydermatis // Ars. Vet. - 4, 1988. - P. 63-68.
11. Mason K., Evans A. Dermatitis associated with Malassezia pachydermatis in 11 dogs // J. Anim. Hosp. Assoc. - 27, 1991. . - P. 13-20.
12. Gaitanis G., Magiatis P. , Hantschke M., Bassukas I., Velegrakid A. The Malassezia Genus in Skin and Systemic Diseases // Clinical Microbiology Reviews. - 25 (1), 2012. - P. 106-141.
13. David M., Gabriel M., Kopecka M. Unusual ultrastructural characteristics of the yeast Malassezia pachydermatis // Scripta medica (Brno). - 76 (3), 2003. - Р. 173-186.
14. Guillot J., Bond R. Malassezia pachydermatis: a review // Medical Mycology. - 37, 1999. - P. 295-306.
15. Benham R. The cultural characteristics of Pityrosporum ovale – a lipophilic fungus // J. Invest. Dermatol. - 2, 1939. - Р. 187-203.
16. Lorenzini R., de Bernardis F. Studies on the isolation, growth and maintenance of Malassezia pachydermatis // Mycopathologia. - 99 (2), 1987. - P. 129-131.
17. Marcon M., Powell D. Human infections due to Malassezia // Clin. Microbiol. Rev. -5(2), 1992. - Р. 101-119.
18. Cafarcia C. Otranto D. Association between phospholipase production by Malassezia pachydermatis and skin lesions // J. clin. Microbiol. - 42(10), 2004 - Р. 4868-4869.
19. Coutinho S., Paula C. Proteinase, phospholipase, hyaluronidase and chondroitin-sulphatase production by Ma-lassezia pachydermatis // Med. Mycol. - 38(1), 2000. -Р. 73-76.
20. Chowdhary A., Randhawa H., Sharma S., Brandt M., Kumar S. Malassezia furfur in a case of onychomycosis: colonizer or etiologic agent? // Med. Mycol. - 43(1), 2005. – Р. 87-90.
21. Bhattacharyya T., Edward M., Cordery C. Colonization of living skin equivalents by Malassezia furfur // Medical Mycology. – 36, 1998. - P. 15-19.
22. Baroni A., Perfetto B., Paoletti I., Ruocco E., Canozo N., Orlando M., Buommino E. Malassezia furfur invasiveness in a keratinocyte cell line (HaCat): effects on cytoskeleton and on adhesion molecule and cytokine expression // Arch Dermatol Res. - 293 (8), 2001 - Р.
414-419.
23. Ashbee H., Evans E. Immunology of diseases associated with Malassezia species // Clin. Microbiol. Rev. – 15 (1), 2002. - Р. 21-57.
24. Tanaka R., Nishimura K., Kamei K., Murayama S. Assimilation test of Malassezia furfur isolated from the environment // Nippon Ishinkin Gakkai Zasshi. - 42(3), 2001. - Р. 123-126.
25. Bond R., Saijonmaa-Koulumies L., Lloyd D. Population sizes and frequency of Malassezia pachydermatis at skin and mucosal sites on healthy dogs // J. small anim. Practice. -36(4), 1995. - P. 147-150.
26. Mayser P., Imkampe A., Winkeler M. Growth requirements and nitrogen metabolism of Malassezia furfur // Arch. Dermatol. Res. – 290, 1998. – Р. 277-282.
27. Арзуманян В.Г., Мокроносова М.А., Гервазиева В.Б. Дрожжеподобные грибы рода Malassezia (Pityrosporum) // Вестник Российской Академии медицинских наук. Вып. 5, 1998. С. 44-47.
28. Foster A., Foil C. BSAVA manual of small animal dermatology // British small animal veterinary association. – Quedgeley, England, 2003. - Р. 175-180.
29. Raabe P., Mayser P., Weiss R. Demonstration of Malassezia furfur and M. sympodialis together with M. pachydermatis in veterinary specimens // Mycoses. – 41 (11-12), 1998. - Р. 493-500.
30. Reme C. Factors associated with Malassezia colonization in the ear canals of healthy dogs // Veterinary Dermatology. – 15 (1), 2004. – P. 11-15.
31. Colombo S., Nardoni S., Cornegliani L., Mancianti F. Prevalence of Malassezia spp. yeasts in feline nail folds: a cytological and mycological study // Vet. Dermatol. -18 (4), 2007. - Р. 278-283.
32. Bond R., Anthony R., Dodd M. Isolation of Malassezia sympodialis from feline skin // J. med. Vet. Mycol. - 34 (2), 1996. - P. 145-147.
33. Cafarchia C. Occurence and population size of Malassezia spp. in the external ear canal of dogs and cats both healthy and with otitis. Mycopathol. -160 (2), -Р. 143-149.
34. Guillot J., Chermette R., Gueho E. Prevalence du genre Malassezia chez les mammiferes // J. Mycol. Med. - 4, 1994. - Р. 72-79.
35. Crespo M., Abarca M., Cabanes F. Occurrence of Malassezia spp. in horses and domestic ruminants // Mycoses. - 45 (7-8), 2002. - P. 333-337.
36. Garau M., del Palacio A., Garc a J. Prevalence of Ma-lassezia spp. in healthy pigs // Mycoses. - 48 (1), 2005. - Р. 17-20.
37. Nakagaki K., Hata K., Iwata E. Malassezia pachydermatis isolated from a south american sea lion (Otaria by-ronia) with dermatitis // J. vet. Med. Sci.- 62 (8), 2000. - Р. 901-903.
38. Midgley G. The lipophilic yeasts: state of the art and prospects // Med. Mycol. - 38 (suppl. 1), 2000. - P. 9-16.
39. Nakabayashi A., Sei Y., Guillot J. Identification of Malassezia species isolated from patients with seborrhoeic dermatitis, atopic dermatitis, pityriasis versicolor and normal subjects // Mwd. Mycol.- 38 (5), 2000. - P. 337-341.
40. Erchiga L., Crespo V., Deglabo V. Malassezia species in skin diseases // Curr. Opin. In skin dis. - 15 (2), 2002. - P. 133-142.
41. Tragiannidis A., Bisping G., Koehler G., Groll A. Minireview: Malassezia infections in immunocompromised patients // Mycoses. - 53 (3), 2010. - P. 187-195.
42. Akerstedt J., Vollset I. Malassezia pachydermatis with special reference to canine skin disease // Br. Vet. J. – 152 (3), 1996. - Р. 269-281.
43. Morris D. Malassezia dermatitis: an opportunist attacks and host responds // The American College of Veterinary Dermatology (ACVD) Resident review. - 2003. - 10 p.
44. Fraser G. Pityrosporum pachydermatis Weidman of canine origin // Trans. Br. Mycol. Soc. - 44, 1961. - P. 441448.
45. Bond R. Pathogenesis of Malassezia dermatitis //Advances in Vet. Dermatol. - 4, 2002. - P. 69-75.
46. Ершов П. Этиологическая значимость дрожжевых грибов рода Malassezia при кожных заболеваниях животных // Дисс.канд. вет. наук. Москва, 2008. - 162 стр.
47. Феофилова Е. Межклеточные взаимодействия в микробных популяциях как основа современной биотехнологии // В кн.: Успехи медицинской микологии. - М.: Национальная академия микологии, 2005, т. 5, с. 231-232.
48. Cannizzo F., Eraso E., Ezkurra P., Villar-Vidal M., Bollo E., Castell G., Caba es F., Vidotto V., Quind s G. Biofilm development by clinical isolates of Malassezia pachydermatis // Med. Mycol. - 45 (4), 2007. - Р. 357-361.
49. Faergemann J. The role of the Malassezia yeasts in skin diseases // Mikol. Lek. - 11 (2), 2004. - P. 129-132.
50. Kumar A., Singh K., Sharma A. Prevalence of Malassezia pachydermatis and other organisms in healthy and infected dogs ears // Israel journal of vet. Med. - 57 (4), 2002. - P. 37-40.
51. Breitwieser F. Results of bacteriologic and mycologic investigations of otitis media in dogs // Tierarztl Prax. - 25 (3), 1997. - Р. 257-260.
52. Ginel P., Lucena R., Rodriguez J., Ortega J. A semiquantitative cytological evaluation of normal and pathological samples from the external ear canal of dogs and cats // Vet. Dermatol. - 13 (3), 2002. - Р. 151-156.
53. Bond R., Patterson-Kane J., Lloyd D. Clinical, histopat-hological and immunological effects of exposure of canine skin to Malassezia pachydermatis // Med. Mycol. - 42 (2), 2004. - Р. 165-175.
54. Chen T., Halliwell R., Hill P. IgG responses to Malassezia pachydermatis antigens in atopic and normal dogs // In: Thoday KL, Foil CS, Bond R, editors. Advances in Veterinary Dermatology. - Vol. 4. - Oxford, England: Blackwell Science, 2002. - P. 202-209.
55. Nuttall T., Halliwell R. Serum antibodies to Malassezia yeasts in canine atopic dermatitis // Vet. Dermatol. - 12, 2001. - P. 327-332.
56. Nagata M., Ishida T. Cutaneous reactivity to Malassezia pachydermatis in dogs with seborrheic dermatitis // In: Proceedings of AAVD/ACVD annual meeting, Santa Fe. - 1995. - P. 11.
57. Bond R., Ferguson E., Curtis C. Factors associated with elevated cutaneous Malassezia pachydermatis populations in dogs with pruritic skin disease // J. small anim. Pract. - 37 (3), 1996. - P. 103-107.
58. Sierra P., Guillot J., Jacob H. Fungal flora on cutaneous and mucosal surfaces of cats infected with feline immunodeficiency virus or feline leukemia virus // Am. J. Vet. Res. - 61 (2), 2000. - P. 158-161.
59. Radi Z. Outbreak of sarcoptic mange and malassezia-sis in rabbits (Oryctolagus cuniculus) // Comp. Med. – 54 (4), 2004. - Р. 434-437.
60. Цыганко А. Болезни ушной раковины и наружного уха. - Бордотекий дог.- 2 (4), 2003. - с. 7-10.
61. Fernandez G., Barboza G., Villalobos A. Isolation and identification of microorganisms present in 53 dogs suffering otitis externa // Revista Cientifica. - 16 (1), 2006. – Р 23-30.
62. Ahman S., Perrins N., Bond R .Carriage of Malassezia spp. yeasts in healthy and seborrhoeic Devon Rex cats // Med. Mycol. - 45 (5), 2007. - Р. 449-455.
63. Duarte E., Batista R., Hahn R. Factors associated with the prevalence of Malassezia species in the external ears of cattle from the state of Minas Gerais, Brazil //Med. Mycol.- 41 (2), 2003. - P. 137-142.
64. Saridomichelakis M., Koutinas A., Gioulekas D. Canine atopic dermatitis in Greece: clinical observations and the prevalence of positive intradermal testreactions in 91 spontaneous cases // Vet. Immunol. Immunopathol. - 69, 1999. - P. 61-73.



Назад в раздел














